How do brains turn environmental inputs into motor outputs?
This question, known as the “black box” problem, has left neuroscientists scratching their heads for decades. On Sept. 5, Amanda Ray, a graduate student in the Gordus Lab at Hopkins, presented her dissertation defense, titled “Understanding How Sensory and Motor Information are Processed in a Neuron of Caenorhabditis elegans.” Her talk suggested that the answer to this question may not lie in the human brain but instead in a little worm by the name of Caenorhabditis elegans (C. elegans).
Ray started off the talk by walking the audience through the key set of neurons in any nervous system: sensory neurons (which detect environmental cues), interneurons (which process and relay those signals) and motor neurons (which tell muscles and glands what to do). Our brains contain billions of these neurons, which communicate with each other, allowing for everything from feeling the slightest of touches to high-level computation.
While it would be nice to map all those connections between neurons to create a connectome, it is unfortunately very challenging. Microscale connectomes are 3D plots of neural structure that allow researchers to connect structure to behavior, but they require intensive electron microscopy. Electron microscopy utilizes high-speed focused electron beams to create magnified images of small specimens that otherwise could not be imaged with a light microscope. Considering that imaging one cubic millimeter of the human brain generates about 1.4 petabytes of data through the examination of 57,000 cells and 150 million synapses, developing a connectome for the brain would be incredibly challenging, requiring an alternate solution.
That’s why neuroscientists like Ray often start small, looking at animals such as mice and fruit flies for modeling. But what made C. elegans the right choice for her work?
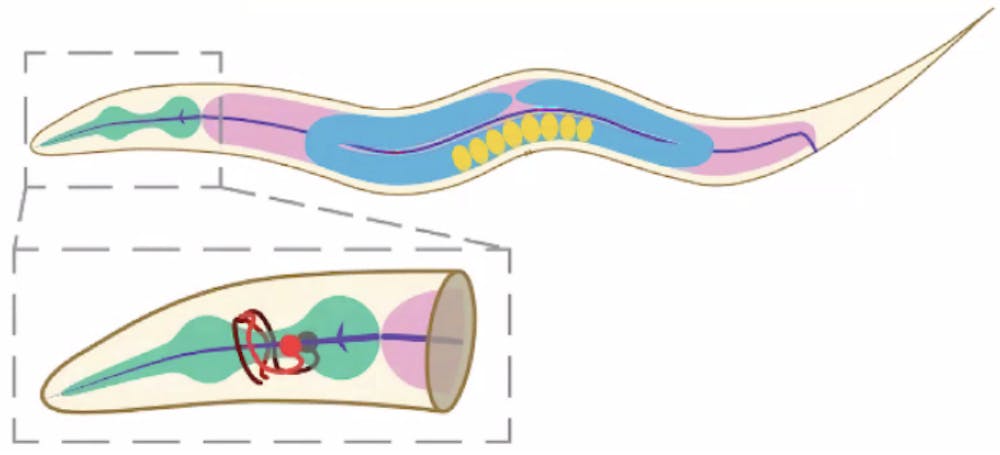
C. elegans has only 302 neurons. It’s transparent and self-fertilizes, which makes researchers’ lives a lot easier.
“Having genetically identical offspring allows developmental and neuronal wiring to also be identical, which is helpful when trying to map neural pathways and tracking neuronal activity of individuals,” Ray said.
And despite its simple neural system, C. elegans performs behaviors such as seeking odors or fleeing from high salt concentrations, proving that these worms possess small but complex displays of sensory-motor decision-making. It should then come as no surprise that scientists have known the entire connectivity of C. elegans since 1986, almost 40 years ago, attesting to the utility of the small yet mighty worm in neuroscience research.
So now that her model organism had been selected, the next step was to find an appropriate neuron to analyze input and output for.
“Modeling helps us fill in the blanks for what goes on in the human brain, illuminating the black box,” Ray explained.
Effectively, she needed interneurons whose inputs and outputs could be clearly tracked. In C. elegans, AVA drives backward movement, RIM modulates AVA’s activity and AIB sits between sensory inputs and these motor commands. Because their wiring and roles are already mapped, they’re ideal for studying how a single neuron integrates sensory and motor signals. Earlier work in the Gordus Lab had shown that motor signals from neurons such as AVA and RIM strongly influence AIB, making AIB a promising neuron for understanding how motor input shapes sensory processing.
Now if neurons were simple calculators, AIB would just sum its inputs and spit out a number that served as an output. Ray wasn’t sure if that was the case and posed three defining questions:
- How does AIB integrate sensorimotor inputs with a small, defined network?
- How does that integration change when one of the inputs is missing?
- How is AIB integrating inputs in the context of the whole brain?
Ray found in her research, through calcium imaging and optogenetics (a technology that captures how neurons light up while stimulating them with flashes of light), AIB is not a basic summation machine when it comes to processing motor-sensory signals.
First, modulating motor input shaped how sensory signals were transmitted through the AIB neuron, meaning that sensory signals don’t possess a fixed effect. The influence of sensory signals depended on motor signalling. When the motor circuit is in one state, sensory inputs are amplified; in another state, they’re muted.
Second, removing motor input changed AIB’s responses to incoming sensory stimuli. Sensory input responses would shrink or shift, implying a relation between how motor and sensory inputs are processed by the AIB neuron.
This led Ray to the conclusion that integrating sensory information in the brain is context-dependent. Even a single worm neuron can “think” about what the body is doing or sensing before deciding how much weight to give incoming sensory data. The motor-dependent model effectively outperformed the summation model.
Ray’s results fit what other worm interneurons revealed. AVA is an interneuron linked to reversal movements, allowing the worm to reverse direction. Another interneuron, RIM, is more complex. RIM can both trigger reversals via fast electrical synapses to AVA and simultaneously hold those reversals in check through slower chemical signals using glutamate and a chloride channel called AVR-14. When RIM is removed from the neural circuit, AVA neurons fire more often but in smaller bursts, and the worm ends up reversing frequently but weakly.
All of this supported Ray’s main finding: sensory input in AIB isn’t processed in a vacuum. The neuron “knows” the worm’s current motor state and changes how it weights incoming sensory data accordingly. Neurons such as RIM show that a single cell can both promote and suppress the same behavior depending on which pathway it is using. Even with a complete wiring diagram, the computation only becomes clear when you watch neurons in action, not just when you trace their connections.
Because of C. elegans, scientists can now better model and describe real computations, such as context-dependent filtering and signal modulation, inside a fully mapped nervous system. And if a 302-neuron worm does this, imagine what’s going on inside our 86-billion-neuron brains.
Preprint of Ray’s dissertation can be accessed on the National Library of Medicine (NIH) website.